A brief history of MEDUSA
In 2004-2005, when NOC had transitioned from the in-house OCCAM general circulation model (GCM) to the pan-European NEMO GCM, there was a gap in the availability of a local marine BGC model. The PISCES and LOBSTER models were available as standard in NEMO, but were judged to be respectively overly and insufficiently complex. The OCCAM’s own (NOC-developed) ecosystem model (bio-OCCAM) was also deemed too simplistic. Against this backdrop, a new model – MEDUSA – was developed to fill this gap.
At this time, MEDUSA was developed during a period of close collaboration between NOC subgroups, including OBE (now OBG), OOC (now MPOC) and OMF (now MSM). It was supported by the regional observational and 3D modelling campaigns of these groups, with collaborative development for three key locations: the Northeast Atlantic, the Northern Atlantic and the Southern Ocean (specifically the Crozet Archipelago). Key contributors in the framing of MEDUSA – in terms of its scope, capabilities and science content – were Katya Popova and Tom Anderson (MSM), Mike Fasham, Raymond Pollard, John Allen, Richard Lampitt, Mike Lucas and Richard Sanders (OBG) and Meric Srokosz (MPOC).
A key focus of MEDUSA was on the carbon cycle in relation to export production and surface-to-deep ocean connectivity. This topic was one of long-term interest within NOC, and continues to be a significant driver of deep oceans observational science. This is largely because of the role (and potential change of this into the future) that marine biology plays in the ocean’s carbon cycle. Theoretical analyses of this cycle suggest that, due to marine biology, atmospheric carbon dioxide (CO2) is at noticeably lower concentrations than would otherwise be expected, and that this is mediated by export production, a complex process involving interactions between many biological actors.
The structure of MEDUSA was created “de novo” but was based loosely on earlier developments at NOC driven by Mike Fasham. Hitherto, three-dimensional GCM work at NOC had focused on nutrient-phytoplankton-zooplankton-detritus models (or subtle variants: e.g. in OCCAM Popova et al., 2006; Yool et al., 2007). However, developments in our understanding of the role of diatoms, the silicon and iron cycles, and size-based differential grazing pressure pointed towards the use of a more sophisticated ecology. Mike Fasham, in conjunction with Ian Totterdell, Katya Popova and Tom Anderson, explored this, and MEDUSA drew from this stream of research within NOC (Popova et al., 2002a,b,2006a,b,2007) with extensive validation on the basis of observational programs conducted by OBE. In parallel, the UK Meteorology Office (UKMO) followed a similar, but distinct, approach with their evolution of their HadOCC model (Palmer & Totterdell, 2001) into Diat-HadOCC (Halloran et al., 2010; Totterdell, 2019).
The processes represented in MEDUSA, and their parameterisation, was drawn from a mixture of extant NOC models plus literature best. In the case of the diatoms and the silicon cycle, work by Phillipe Pondaven and Mathieu Mongin informed the approach used here (Mongin et al., 2006), and MEDUSA includes a dynamic Si:N ratio. In the case of the iron cycle, the simplified (1 tracer, fixed stoichiometry) approach of Mick Follows and Payal Parekh was adopted (Parekh et al., 2005). And in the case of export production, a formulation of Robert Armstrong’s ballast model was adopted (Armstrong et al., 2002) and developed into the fast-sinking particles of MEDUSA’s two size-class model of sinking detritus (derived, in part, from the modelling work of John Dunne; Dunne et al., 2007).
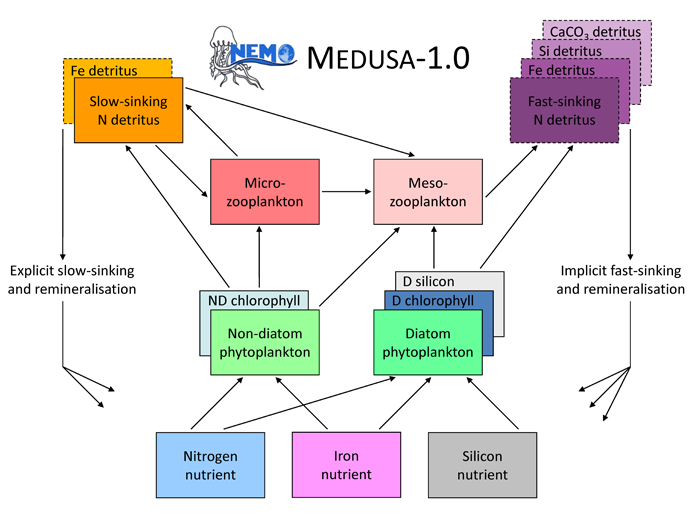
During Oceans 2025, and despite the carbon focus of this programme, MEDUSA-1 was developed as a nitrogen-silicon-iron model, in which carbon fluxes were inferred stoichiometrically. This version of MEDUSA was documented and evaluated in Yool et al. (2011), Popova et al., (2010, 2012) and its schematic is shown below.
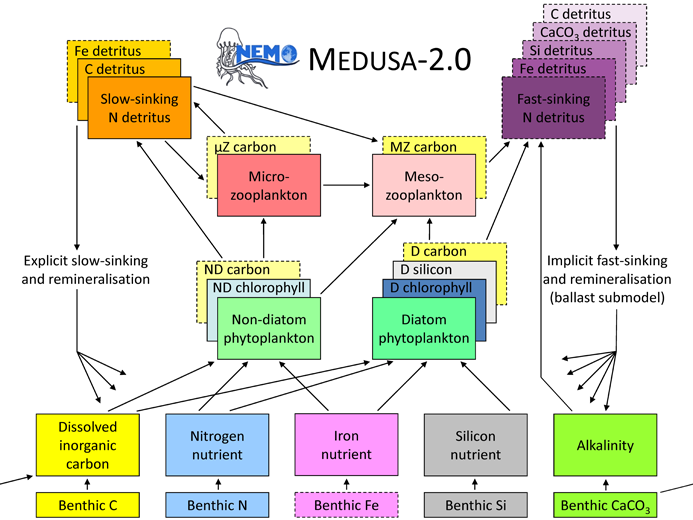
Subsequently, and related to the Regional Ocean Acidification Modelling (ROAM) project of the UK’s Ocean Acidification Research Programme (UKOARP), MEDUSA-2 was extended to include the cycles of carbon, alkalinity and oxygen. At this point, and to the present day, while phytoplankton and zooplankton in MEDUSA were permitted to have different stoichiometry, this was still assumed to be fixed, with only detrital particulate organic carbon (POC) having variable C:N stoichiometry. This version of MEDUSA was documented and evaluated in Yool et al. (2013), and used actively for Yool et al. (2013b), Popova et al. (2013, 2014,2016) and Yool et al. (2015). Its schematic is shown below.
As part of ROAM, a ground-breaking future ocean projection was performed using MEDUSA at 1/4-degree resolution – a step above of typical 1-degree resolution of Earth system models (ESMs). This run provided a unique capability to analyse impacts of future changes in the boundary currents and upwelling systems on ocean primary production. This run was applied to a range of topics and geographical regions across a range of publications, including: Yool et al., 2015, Popova et al. (2016), Jacobs et al. (2022), Asdar et al. (2022).
During a subsequent project, iMarNet (2012-2014), MEDUSA-2 was evaluated for its potential as the marine biogeochemistry submodel of the UK’s Earth system model (ESM) then beginning development for CMIP6. In iMarNet, MEDUSA-2 “competed” against 5 other UK marine biogeochemistry models in an exercise to evaluate their overall scientific performance and suitability for use. The other models considered included HadOCC (NOC, UKMO), Diat-HadOCC (UKMO), PlankTOM-6 (UEA), PlankTOM-10 (UEA) and ERSEM (PML), and spanned a large range of complexity from HadOCC’s simple, NPZD framework (7 state variables), out to ERSEM’s multiple plankton types structure (57 state variables). A range of biogeochemical evaluation targets were used, but key assessment targets in iMarNet were the realism of the carbon cycle and the computational cost of each model. Biogeochemical performance varied across the models, with no one model consistently superior across all targets. Meanwhile, model compute cost was found to scale linearly with tracer burden, such that HadOCC slightly less than doubled total model cost, while ERSEM increased it by more than a factor of 10. However, the absence of iron and silicon cycles in HadOCC ultimately precluded its use and, sitting in “second place”, MEDUSA-2 was selected as the marine biogeochemistry component for the joint NERC / UKMO Earth system model, UKESM1. The evaluation in iMarNet is documented in Kwiatkowski et al. (2014).
At the same time, MEDUSA was incorporated into global hindcast runs covering the period of 1990-2015, being the first global coupled physical and biogeochemical model run globally at 1/12-degree resolution. This run underpinned a number of regionally focused studies addressing dynamics of the shelf upwelling regions (Jacobs et al., 2020a,b; 2022; Jebri et al., 2022) as well as a series of policy briefs dedicated to the management of the Tanzanian and Kenyan fisheries: [https://solstice-wio.org/outputs/policy-briefs].
After its selection for use in UKESM1, MEDUSA-2 scientific development was largely frozen to ensure traceability and to facilitate and expedite its use within the ESM. Developments that did take place included the replacement of MEDUSA-2’s carbonate chemistry (to OMIP's preferred MOCSY; Orr et al., 2015), and inclusion of a DMS parameterisation to permit atmospheric sulphur cycle feedbacks (Anderson et al. 2001). Technical model improvements continued during UKESM1’s development to accommodate up-to-date NEMO revisions (moving from NEMO v3.2 to v3.6), to improve diagnostic outputting, to restructure and modernise its code, and to include a number of bug fixes, including to model conservation. This work was mostly completed by Julien Palmieri, who joined MSM in 2014 for this specific task (and leading into UKESM2). To distinguish it, the version of MEDUSA used in UKESM1 is formally described as MEDUSA-2.1 because of these and other developments.
From 2016-2018, UKESM1 was spun-up, first in ocean-only mode, before subsequently transitioning to the fully-coupled model. The ocean phase of this process involved a NEMO-MEDUSA simulation of 4800 years, an unprecedented duration in NOC’s experience. Physical and biogeochemical trends during this simulation, together with an analysis of the final pre-industrial state of the model (all components) form the basis of the UKESM1 paper, Yool et al. (2020).
During 2018 and continuing through 2019, UKESM1 (including MEDUSA-2.1) performed CMIP6 simulations. Evaluation found good traceability for MEDUSA-2 between previous historical period simulations (cf. UKOARP) and current UKESM1 simulations. A formal analysis of the ocean system, including MEDUSA-2.1, has been published by NOC (Yool et al., 2021).
As part of this CMIP6 activity, specifically future climate projections, MEDUSA (as part of UKESM1) was widely used in the Working Group 1 (WG1) component of the IPCC’s 6th Assessment Report (AR6). WG1 addreses the most up-to-date physical understanding of the climate system and climate change, bringing together the latest advances in climate science. UKESM1 projections are referenced in the IPCC AR6 WGI report more than 100 times, including in 29 Figures and 7 tables. 8 papers involving MEDUSA as part of the UKESM1 are cited in the report, with 5 involving NOC co-authors. MEDUSA publications (Yool et al., 2013, 2015, 2017) were additionally cited in the Working Group 2 (WG2) component of AR6, which addresses impacts, adaptation and vulnerability. Here, MEDUSA and its benthic submodel, BORIS, were used as key evidence for the future of seafloor ecosystems (Figure 3.21j,k,l, from the WG2 report was re-calculated using BORIS forced with UKESM1 output, following Yool et al., 2017).
Currently, MEDUSA is being prepared for its inclusion in the UKESM2 model, and will take part in CMIP7 simulations expected to occur after 2025.
During the ongoing Climate-Linked Atlantic Sector Science (CLASS) project, MEDUSA is being extended to include: 1. Dynamic phytoplankton C:N stoichiometry; 2. Temperature-dependent zooplankton ecophysiology; 3. Open nitrogen cycle, with riverine inputs and nitrogen-fixation.
CLASS has also driven the production of a new suite of future-forced runs to update that undertaken during the ROAM project. Simulations have been performed at 1-, 1/4- and 1/12-degree resolution for the full Historical period (1850-2014) and into CMIP6’s Scenario period (2015-2100). These simulations used UKESM1.1 output, running under the high emissions SSP370 Scenario (“Regional Rivalry”). These runs provide a cutting-edge resource for analysis of future projections at resolutions beyond those typical for global-scale models.

(top row) Mike Fasham, Tom Anderson, Katya Popova, Ian Totterdell, Andrew Yool, and Julien Palmieri; (bottom row) Raymond Pollard, John Allen, Richard Sanders, Meric Srokosz, Richard Lampitt, and Mike Lucas.
- Anderson, T. R., Spall, S. A., Yool, A., Cipollini, P., Challenor, P.G., and Fasham, M. J. R.: Global fields of sea surface dimethylsulphide predicted from chlorophyll, nutrients and light, J. Marine Syst., 30, 1–20, 2001.
- Armstrong, R. A., Lee, C., Hedges, J. I., Honjo, S., and Wakeham, S. G.: A new, mechanistic model for organic carbon fluxes in the ocean: based on the quantitative association of POC with ballast minerals, Deep-Sea Res. Pt. II, 49, 219–236, 2002.
- Asdar, S., Jacobs, Z.L., Popova, E., Noyon, M., Sauer, W., and Roberts, M.J.: Projected climate change impacts on the ecosystems of the Agulhas Bank, South Africa, Deep Sea Research Part II: Topical Studies in Oceanography, 105092, https://doi.org/10.1016/j.dsr2.2022.105092, 2022.
- Dunne, J. P., Sarmiento, J. L., and Gnanadesikan, A.: A synthesis of global particle export from the surface ocean and cycling through the ocean interior and on the seafloor, Global Biogeochem. Cy., 21, GB4006, doi:10.1029/2006GB002907, 2007.
- Halloran, P. R., Bell, T. G., and Totterdell, I. J.: Can we trust empirical marine DMS parameterisations within projections of future climate?, Biogeosciences, 7, 1645–1656, https://doi.org/10.5194/bg-7-1645-2010, 2010.
- Jacobs, Z.L., Jebri, F., Srokosz, M., Raitsos, D.E., Painter, S.C., Nencioli, F., Osuka, K., Samoilys, M., Sauer, W., Roberts, M., Taylor, S.F.W., Scott, L., Kizenga, H., and Popova E: A Major Ecosystem Shift in Coastal East African Waters During the 1997/98 Super El Niño as Detected Using Remote Sensing Data. Remote Sensing, 12, 3127, https://doi.org/10.3390/rs12193127, 2020a.
- Jacobs, Z.L., Jebri, F., Raitsos, D.E., Popova, E., Srokosz, M., Painter, S.C., Nencioli, M., Roberts, J., Kamau, M., Palmer, J., and Wihsgott: Shelf-break upwelling and productivity over the North Kenya Banks: The importance of large-scale ocean dynamics, Journal of Geophysical Research, 125, e2019JC015519, https://doi.org/10.1029/2019JC015519, 2020b.
- Jacobs, Z.L., Yool, A., Jebri, F., Srokosz, M., van Gennip, S., Kelly, S.J., Roberts, M., Sauer, W., Queirós, A.M., Osuka, K.E., Samoilys, M., Becker, A.E., and Popova, E.: Key climate change stressors of marine ecosystems along the path of the east african coastal current, Ocean & Coastal Management, 208, 105627, https://doi.org/10.1016/j.ocecoaman.2021.105627, 2021.
- Jebri, F., Raitsos, D.E., Gittings, J.A., Jacobs, Z.L., Srokosz, M., Gornall, J., Sauer, W.H.H., Roberts, M.J., Popova, E.: Unravelling links between squid catch variations and biophysical mechanisms in South African waters, Deep Sea Research Part II: Topical Studies in Oceanography, 196, 105028, https://doi.org/10.1016/j.dsr2.2022.105028, 2022.
- Kwiatkowski, L., Yool, A., Allen, J.I., Anderson, T.R., Barciela, R., Buitenshuis, E.T., Butenschon, M., Enright, C., Halloran, P.R., Le Quere, C., de Mora, L., Racault, M.F., Sinha, B., Totterdell, I.J., Cox, P.M. (2014). iMarNet: an ocean biogeochemistry model intercomparison project within a common physical ocean modelling framework. Biogeosciences 11, 7291-7304, doi: 10.5194/bg-11-7291-2014.
- Mongin, M., Nelson, D. M., Pondaven, P., and Tréguer, P.: Simulation of upper-ocean biogeochemistry with a flexible-composition phytoplankton model: C, N and Si cycling and Fe limitation in the Southern Ocean, Deep-Sea Res. Pt. II, 53, 601–619, 2006.
- Orr, J. C. and Epitalon, J.-M.: Improved routines to model the ocean carbonate system: mocsy 2.0, Geosci. Model Dev., 8, 485–499, doi: 10.5194/gmd-8-485-2015, 2015.
- Palmer, J.R.; Totterdell, I.J.: Production and export in a global ocean ecosystem model. Deep-Sea Research I, 48, 1169-1198, https://doi.org/10.1016/S0967-0637(00)00080-7, 2001.
- Parekh, P., Follows, M. J., and Boyle, E. A.: Decoupling of iron and phosphate in the global ocean, Global Biogeochem. Cy., 19, GB2020, doi:10.1029/2004GB002280, 2005.
- Popova, E.E., Yool, A., Aksenov, Y., Coward, A.C., Anderson, T.R. (2014). Regional variability of acidification in the Arctic: a sea of contrasts. Biogeosciences 11, 293-308, doi: 10.5194/bg-11-293-2014.
- Popova, E., Yool, A., Byfield, V., Cochrane, K., Coward, A.C., Salim, S.S., Gasalla, M.A., Henson, S.A., Hobday, A.J., Pecl, G., Sauer, W., and Roberts, M.: From global to regional and back again: common climate stressors of marine ecosystems relevant for adaptation across five ocean warming hotspots. Global Change Biology, 22, 2038-2053, https://doi.org/10.1111/gcb.13247, 2016.
- Totterdell, I. J.: Description and evaluation of the Diat-HadOCC model v1.0: the ocean biogeochemical component of HadGEM2-ES, Geosci. Model Dev., 12, 4497–4549, https://doi.org/10.5194/gmd-12-4497-2019, 2019.
- Yool, A., Martin, A., Fernández, C. et al.: The significance of nitrification for oceanic new production. Nature 447, 999–1002, https://doi.org/10.1038/nature05885, 2007.
- Yool, A., Popova, E.E. and Anderson, T.R. (2011). MEDUSA-1.0: a new intermediate complexity plankton ecosystem model for the global domain. Geoscientific Model Development 4, 381-417, doi: 10.5194/gmd-4-381-2 011.
- Yool, A., Popova, E.E. and Anderson, T.R. (2013). MEDUSA-2.0: an intermediate complexity biogeochemical model of the marine carbon cycle for climate change and ocean acidification studies. Geoscientific Model Development 6, 1767-1811, doi: 10.5194/gmd-6-1767-2013.
- Yool, A., Popova, E.E., Coward, A.C., Bernie, D. and Anderson, T.R. (2013b). Climate change and ocean acidification impacts on lower trophic levels and the export of organic carbon to the deep ocean. Biogeosciences, 10, 5831-5854, doi: 10.5194/bg-10-5831-2013.
- Yool, A., Popova, E.E. and Coward, A.C. (2015). Future change in ocean productivity: Is the Arctic the new Atlantic? J. Geophys. Res. 120, 7771-7790, doi: 10.1002/2015JC011167.
- Yool, A., Palmiéri, J., Jones, C.G., Sellar, A.A., de Mora, L., Kuhlbrodt, T., Popova, E.E., Mulcahy, J.P., Wiltshire, A., Rumbold, S.T., Stringer, M., Hill, R.S.R., Tang, Y., Walton, J., Blaker, A., Nurser, A.G.J., Coward, A.C., Hirschi, J., Woodward, S., Kelley, D.I., Ellis, R., Rumbold‐Jones, S. (2020). Spin‐up of UK Earth System Model 1 (UKESM1) for CMIP6. Journal of Advances in Modeling Earth Systems, 12, e2019MS001933, doi: 10.1029/2019MS001933.
- Yool, A., Palmiéri, J., Jones, C. G., de Mora, L., Kuhlbrodt, T., Popova, E. E., Nurser, A. J. G., Hirschi, J., Blaker, A. T., Coward, A. C., Blockley, E. W., and Sellar, A. A.: Evaluating the physical and biogeochemical state of the global ocean component of UKESM1 in CMIP6 historical simulations, Geosci. Model Dev., 14, 3437–3472, doi: 10.5194/gmd-14-3437-2021, 2021.