Model description
MEDUSA’s ecosystem is structured as a dual size-class nutrient-phytoplankton-zooplankton-detritus (NPZD) model. Dissolved inorganic nutrients – including nitrogen, silicon, iron and carbon – form the base of the model. Phytoplankton are represented by “small” nanophytoplankton, typically photosynthetic prokaryotes, and “large” microphytoplankton, assumed synonymous with siliceous diatoms. Both phytoplankton groups have a dynamic chlorophyll quota to allow them to photoacclimate across a range of surface, submarine and seasonal light conditions. In turn, MEDUSA’s phytoplankton are consumed by two size classes of heterotrophic grazers, microzooplankton, assumed to be fast-growing single-celled protists, and mesozooplankton, assumed to be multicellular metazoans, such as copepods. Mortality and other loss processes that affect these plankton components results in the production of particles of non-living detrital material that sink into the ocean interior. As with the other components, these are divided into “small”, slow-sinking particles that are predominantly remineralised in the upper euphotic zone, and “large”, fast-sinking particles that form the primary route by which organic material reaches the deep ocean. At the seafloor, MEDUSA resolves 4 reservoirs to temporarily store organic (nitrogen and carbon) and inorganic (opal and CaCO3) material reaching the sediment via both slow- and fast-sinking particles.
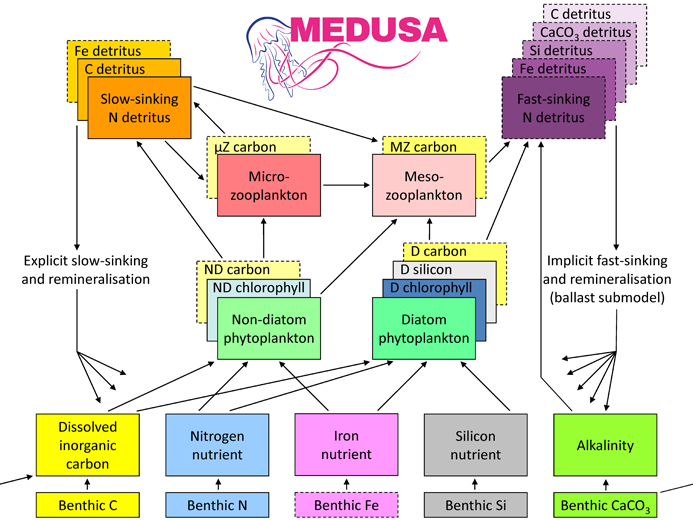
The schematic above presents an outline of MEDUSA's components and the process connections between them. The model includes the biogeochemical cycles of nitrogen, silicon, iron, carbon, alkalinity and oxygen in a dual size class nutrient-phytoplankton-zooplankton-detritus framework. Components with solid borders are those explicitly represented as passive tracers in MEDUSA-2.1, while those with dashed borders are implicit to reduce model cost. These are either linked via rigid stoichiometry to explicit components (e.g. carbon in plankton) or occur only temporarily (e.g. fast-sinking detritus). Oxygen has been omitted from the diagram for simplicity.
The model's nitrogen, silicon and alkalinity cycles are closed and conservative (e.g. no riverine inputs), while the cycles of iron, carbon and oxygen are open. The ocean's iron cycle includes additions from aeolian and benthic sources, and is depleted by scavenging based on local iron availability (and an assumed constant binding ligand concentration). The ocean's carbon cycle exchanges CO2 with the atmosphere based on local carbonate chemistry, atmospheric CO2 concentration and ambient winds. The ocean's oxygen cycle exchanges with the atmosphere (which has an assumed constant oxygen concentration), and dissolved oxygen is additionally created by primary production in the euphotic zone and depleted by remineralisation throughout the ocean. The various elemental cycles include both fixed and variable stoichiometry. Iron is slaved to nitrogen throughout, while nitrogen and carbon have fixed (but different) ratios in phytoplankton and zooplankton, and variable ratios in detritus. Diatom silicon has a variable ratio with nitrogen, dependent on nutrient availability and growth rate. Calcium carbonate is produced at a geographically-variable rate relative to organic carbon according to the ambient calcite saturation state, and consumes both dissolved inorganic carbon (DIC) and alkalinity in a ratio of 1:2 respectively. Oxygen production and consumption reflects the C:N ratio of organic matter produced and consumed.
Yool et al. (2013) extensively describes the structure, differential equations, functional forms and parameterisation of the MEDUSA-2.0 model in an earlier, ocean-only configuration. As part of the development cycle of UKESM1, a number of changes were made to the model, and the resulting version used is denoted as MEDUSA-2.1 for clarity. The specific developments are listed below.
- The carbonate chemistry submodel used in MEDUSA-2 is now the MOCSY-2.0 scheme of Orr et al. (2015).
- Since UKESM1 represents atmospheric chemistry, including elements of the sulphur cycle, MEDUSA now includes the empirical submodel of Anderson et al. (2001) to estimate surface dimethyl sulphide (DMS) concentration and exchange of this to the atmosphere.
- In addition to DMS, the atmospheric chemistry submodel of UKESM1 includes the emission of primary marine organic aerosol (PMOA). This utilises MEDUSA's simulated surface chlorophyll coupled to the PMOA parameterisation of Gantt et al. (2011) and Gantt et al. (2012).
The performance of MEDUSA in UKESM1 is evaluated at length in the assessment of Yool et al. (2021), including against peer CMIP6 models.
BORIS
The BORIS model was developed in NOC to represent the abundance of benthic organisms on the seafloor of, principally, the deep ocean. From observational work, it was known that organism size plays an important role in these communities, potentially more than functional or taxonomic groupings. As size (specifically mass) is readily quantified, and correlates well with metabolism and abundance, it makes for an attractive framework, both practically and theoretically. While debate on the causes and universality of this correlation continues, size-based approaches to communities have proven useful, in both marine and terrestrial settings.
Building from this, BORIS represents the benthic community as a size spectrum of heterotrophic consumers that feed on sinking particles of organic carbon that reach the seafloor. These organisms all consume this organic carbon from a single common reservoir, returning it back to solution as dissolved CO2 through respiration, or back into the reservoir in the case of egestion of waste or mortality. The size spectrum of BORIS is discretised into a number of size classes, and the parameters (growth, respiration and mortality rates) for each class follow a power law relationship based on size (i.e. allometric rates).
References
- Anderson, T. R., Spall, S. A., Yool, A., Cipollini, P., Challenor, P.G., and Fasham, M. J. R.: Global fields of sea surface dimethylsulphide predicted from chlorophyll, nutrients and light, J. Marine Syst., 30, 1–20, doi: 10.1016/S0924-7963(01)00028-8, 2001.
- Gantt, B., Meskhidze, N., Facchini, M. C., Rinaldi, M., Ceburnis, D., and O’Dowd, C. D.: Wind speed dependent size-resolved parameterization for the organic mass fraction of sea spray aerosol, Atmos. Chem. Phys., 11, 8777–8790, doi: 10.5194/acp-11-8777-2011, 2011.
- Gantt, B., Johnson, M. S., Meskhidze, N., Sciare, J., Ovadnevaite, J., Ceburnis, D., and O’Dowd, C. D.: Model evaluation of marine primary organic aerosol emission schemes, Atmos. Chem. Phys., 12, 8553–8566, doi: 10.5194/acp-12-8553-2012, 2012.
- Orr, J. C. and Epitalon, J.-M.: Improved routines to model the ocean carbonate system: mocsy 2.0, Geosci. Model Dev., 8, 485–499, doi: 10.5194/gmd-8-485-2015, 2015.
- Yool, A., Popova, E.E. and Anderson, T.R. (2013). MEDUSA-2.0: an intermediate complexity biogeochemical model of the marine carbon cycle for climate change and ocean acidification studies. Geoscientific Model Development 6, 1767-1811, doi: 10.5194/gmd-6-1767-2013.
- Yool, A., Palmiéri, J., Jones, C. G., de Mora, L., Kuhlbrodt, T., Popova, E. E., Nurser, A. J. G., Hirschi, J., Blaker, A. T., Coward, A. C., Blockley, E. W., and Sellar, A. A.: Evaluating the physical and biogeochemical state of the global ocean component of UKESM1 in CMIP6 historical simulations, Geosci. Model Dev., 14, 3437–3472, doi: 10.5194/gmd-14-3437-2021, 2021.
- Kelly-Gerreyn, B.A., Martin, A.P., Bett, B.J., Anderson, T.R., Kaariainen, J.I., Main, C.E., Marcinko, C.J. and Yool, A.: Benthic biomass size spectra in shelf and deep-sea sediments. Biogeosciences 11, 6401-6416, doi: 10.5194/bg-11-6401-2014, 2014.
- Yool, A., Martin, A.P., Anderson, T.R., Bett, B.J., Jones, D.O.B. and Ruhl, H.A.: Big in the benthos: Future change of seafloor community biomass in a global, body size-resolved model. Glob. Change Biol. 23, 3554-3566, doi: 10.1111/gcb.13680, 2017.
- Kelly-Gerreyn, B.A., Martin, A.P., Bett, B.J., Anderson, T.R., Kaariainen, J.I., Main, C.E., Marcinko, C.J. and Yool, A.: Benthic biomass size spectra in shelf and deep-sea sediments. Biogeosciences 11, 6401-6416, doi: 10.5194/bg-11-6401-2014, 2014.
- Yool, A., Martin, A.P., Anderson, T.R., Bett, B.J., Jones, D.O.B. and Ruhl, H.A.: Big in the benthos: Future change of seafloor community biomass in a global, body size-resolved model. Glob. Change Biol. 23, 3554-3566, doi: 10.1111/gcb.13680, 2017.